Strasznie przepraszam, ale nie mam dziś czasu przerabiać tego. Wklejam więc fragment tłumaczonej przeze mnie książki "Compendium of animal reproduction" PC Nelis, Intervet. Naprawdę, nie dam dziś rady napisac nic sensownego. Mam nadzieję, że to nikogo nie zabije. Latwiej mi będzie odpowiedziec na ewentualne pytania, niż pisac wszystko od nowa. Poza tym, naprawdę nie wiem, jak ująć to w sposob przystępny

No, przepraszam
 Wstęp
Wstęp
U ssaków w procesy rozrodu zaangażowane są dwa systemy regulacyjne. Układ wydzielania wewnętrznego i układ nerwowy razem grają specyficzną rolę. Subtelne współgranie obu układów jest zasadnicze dla łańcuch zdarzeń, który ostatecznie prowadzi do porodu i udanego odchowu zdrowego potomstwa.
Układ nerwowy, układ wydzielania wewnętrznego i mediatory komórkowe
Bodźce z otoczenia są odbierane przez narządy zmysłów i przekazywane do mózgu. Postrzeganie zmysłowe może mieć różny charakter. Oczy (światło lub widok innych zwierząt tego samego gatunku), nos (zapachy płciowe) i zmysł dotyku odbierają bodźce z otoczenia a nerw wzrokowe, nerw węchowy i nerwy czuciowe przesyłają informacje do mózgu. Mózg jest zdolny odczytać informacje i, jeżeli jest to potrzebne, zareagować, przesyłając impuls nerwowy przez włókna nerwowe do narządu docelowego.
Układ hormonalny jest układem regulacyjnym, przesyłającym informacje za pomocą przekaźników chemicznych. System ten regulowany jest przez sprzężenia zwrotne i impulsy z układu nerwowego i innych narządów.
Hormon można określić jako związek chemiczny, wytwarzany w gruczole lub w tkance ciała, wywołujący specyficzną reakcje w tkance wrażliwej na działanie hormonu. Działanie układu wydzielania wewnętrznego można podzielić zależnie od drogi pokonywanej przez hormony do komórek docelowych. W drodze autokrynnej komórka wytwarzająca hormon jest zarazem komórką docelową. W drodze parakrynnej, wytwarzane hormony wpływają na sąsiadujące komórki i narządy. Hormony endokrynne są transportowane przez krew, a w drodze egzokrynnej hormony są wydzielane (w przewodzie pokarmowym, układzie moczowym, na skórze itp.) przed osiągnięciem narządów docelowych. Jednak najwięcej informacji jest przekazywanych drogą endokrynną. W czasie ostatniej dekady badacze poświęcili dużo uwagi funkcjom para- i autokrynnym układu hormonalnego, ale wiele aspektów ich funkcjonowania nadal pozostaje nie odkryte.
Po osiągnięciu komórki docelowej hormon wywołuje reakcję. W tym celu komórka ma receptory specyficzne dla hormonu. Receptor dla hormonu jest unikalną strukturą molekularną na powierzchni lub wewnątrz komórki z wysokim i specyficznym powinowactwem dla określonego hormonu. Po połączeniu z receptorem może zostać odczytana informacja. Ta wiadomość prowadzi do komórkowo specyficznej odpowiedzi, która zasadniczo powoduje aktywacje lub inaktywację enzymów komórki docelowej.
Receptory dla hormonów maja wiec dwie zasadnicze funkcje:
- Rozpoznanie specyficznego hormonu przez komórkę docelową.
- Przetworzenie sygnału hormonalnego na komórkowo specyficzną odpowiedź.
Efekt endokrynnego uwolnienia hormonów może zależeć od wielu okoliczności. Ilość i rodzaj receptorów komórki docelowej nie jest wartością stałą. Tworzenie i degradacja receptorów jest procesem dynamicznym. Zadaniem hormonu w komórce może być indukcja lub degradacja receptorów dla innego przekaźnika. Ponadto, receptory mogą być blokowane przez nadmiar hormonów. Nadmierna stymulacja przez zwykle wysokoefektywną dawkę hormonu, może później nie wywołać więcej efektu. Wiele stanów patologicznych w rozrodzie jest powodowanych zaburzeniami na poziomie receptorowym.
Większość receptorów potrzebuje wtórnego przekaźnika do przekazania wiadomości. Jednym z najlepiej poznanych wtórnych przekaźników jest cAMP.
Sposób połączenia układu nerwowego z układem wydzielania wewnętrznego będzie omówiony w następnym paragrafie.
Regulacja rozrodu u samic
Okresy bez regularnej cyklicznej aktywności (anoestrus) stanowią większą część życia normalnie płodnej samicy. Okres młodzieńczego oraz ciążowego i laktacyjnego anoestrus zajmują dużo więcej czasu niż relatywnie krótkie okresy cyklicznej aktywności.
Jednakże najwięcej uwagi poświęca się okresom cyklicznej aktywności. Jest to okres, w którym człowiek najczęściej ingeruje w procesy rozrodu (rozmnażać/nie rozmnażać; wybór samca/sztuczne zapłodnienie, kontrola rui, wywoływanie owulacji itp.), a także okres, w którym najczęściej występują problemy związane z rozmnażaniem.
Ogólne zasady regulacji hormonalnej rozrodu są takie same dla różnych gatunków, ale występują jednak charakterystyczne różnice pomiędzy gatunkami. Niektóre zwierzęta są poliestralne (krowa, maciora) przez wszystkie lata, niektóre są sezonowo poliestralne (klacz, owca, kotka). Suki są monoestralne. Inne różnice są w sposobie owulacji. U większości zwierząt występuje owulacja spontaniczna, ale u kotki, królicy i wielbłądzicy owulacja jest indukowana przez stymulacje receptorów czuciowych w pochwie i szyjce w trakcie kopulacji. Szczegółowo zagadnienia procesów rozrodu u poszczególnych gatunków są omówione w odpowiednich rozdziałach dotyczących fizjologii rozrodu. W tej części zostaną jedynie przedstawione najważniejsze hormony i tkanki (wydzielnicze jak i docelowe), uczestniczące w procesach rozrodu. Cykl płciowy krowy jest użyty jako przykład do wyjaśnienia funkcji i interakcji różnych narządów i hormonów.
Procesy rozrodu u ssaków są regulowane kompleksowo, i tylko częściowo poznane, przez kaskadę impulsów z ośrodkowego układu nerwowego, tkanki wydzielnicze, tkanki docelowe i niektóre hormony.
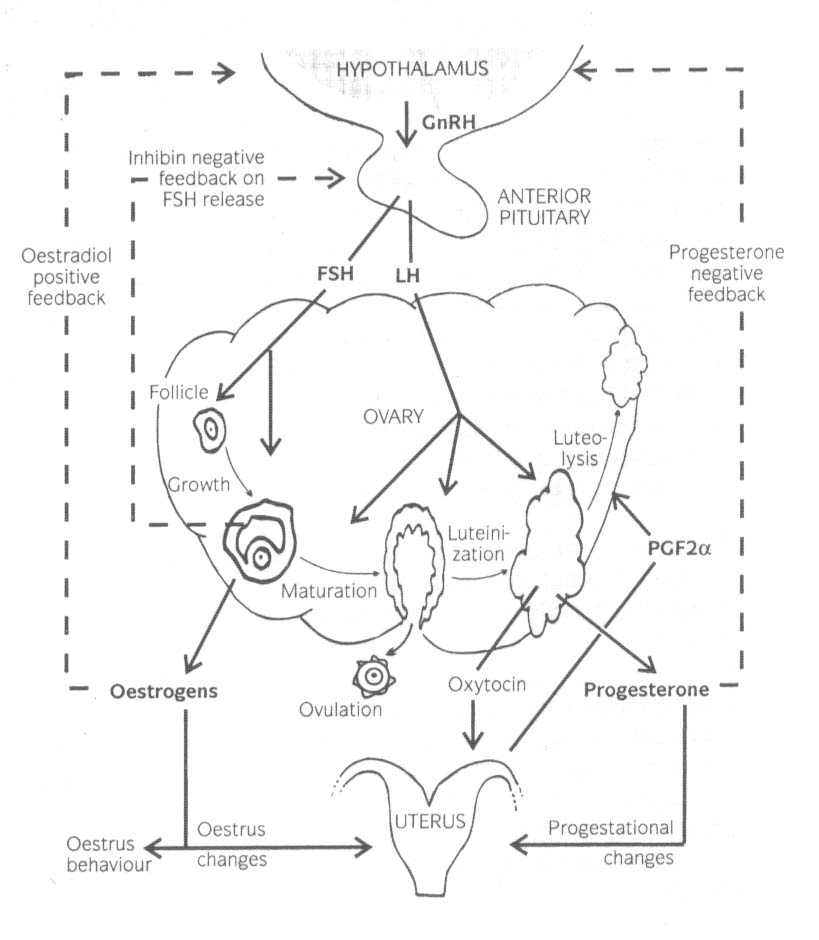
Rycina przedstawia schematycznie najważniejsze narządy i hormony zaangażowane w procesy rozrodu u samicy, uwzględniając niektóre ich funkcje i interakcje.
Ośrodkowy układ nerwowy odbiera informacje z otoczenia zwierzęcia (sygnały zewnętrzne: wzrokowe, węchowe, słuchowe, czuciowe) i przesyła je, jeżeli mają one znaczenie dla reprodukcji, do gonad za pomocą osi podwzgórzowo-przysadkowo-gonadowej. Podwzgórze i przysadka są strukturami ściśle związanymi z brzuszną częścią mózgu. Obie struktury są nie tylko producentami hormonów, ale także narządami docelowymi, tworzącymi homeostatyczny system sprzężeń zwrotnych. Poprzez ten mechanizm sprzężeń zwrotnych większość hormonów reguluje swoje własne wytwarzanie.
W podwzgórzu neurony endokrynowe produkują, po pobudzeniu z osrodkowego układu nerwowego, gonadoliberynę (GnRH). GnRH jest transportowane poprzez podwzgórzowo-przysadkowy układ wrotny do przedniego płata przysadki. Tu stymuluje komórki gonadotropowe przysadki do sekrecji hormonu follikulotropowego (FSH) i hormonu luteinizującego (LH). GnRH, FSH i LH są uwalniane pulsacyjnie.
FSH stymuluje rozwój pęcherzyków jajnikowych. W osłonce wewnętrznej LH stymuluje syntezę androstendionu z cholesterolu. Androstendion jest przekształcany do testosteronu, który w komórkach warstwy ziarnistej jest aromatyzowany do 17-beta-estradiolu pod wpływem FSH. Estradiol wykazuje sprzężenie zwrotne dodatnie podwzgórze i przysadkę. Zwiększa częstotliwość pulsów GnRH. Powyżej pewnego poziomu estradiolu podwzgórze odpowiada wylewem GnRH. Ten wylew GnRH wywołuje wylew LH, co zapoczątkowuje owulację. Innym ważnym efektem działania estradiolu jest wywoływanie oznak rui. Ruja może zostać opisana jako behawioralne i fizyczne objawy, sygnalizujące innym zwierzętom, ze samica jest w okresie płodnym i dopuści samca do krycia.
Komórki warstwy ziarnistej produkują także inhibinę. Nie wszystkie efekty działania tego hormonu są poznane, ale zawdzięcza on swoją nazwę ujemnemu sprzężeniu zwrotnemu na uwalnianie FSH z przysadki, przez co kontroluje rozwój pęcherzyków.
Po owulacji pozostałości pęcherzyka są przekształcane w ciałko żółte pod wpływem LH. Dziura w pęcherzyku jest wypełnione krwią z naczyń, a warstwa ziarnista powiększa się. Ciałko żółta jest głównie narządem wydzielniczym, produkującym progesteron i oksytocynę. Progesteron jest niezbędny dla prawidłowego przebiegu cyklu płciowego u krowy, a po zapłodnieniu jest podstawowym hormonem zapewniającym utrzymanie ciąży. Zmniejsza on pulsy GnRH i w ten sposób zapobiega ponownej owulacji. Ponadto, przygotowuje endometrium do zagnieżdżenia rozwijającego się zarodka i zapobiega niekontrolowanym przeciwdziałaniom ze strony ściany macicy względem płodu.
Jeżeli komórka jajowa uwolniona z pęcherzyka w czasie owulacji nie zostanie zapłodniona, zwierzę nie otrzyma sygnału ciąży od zarodka. Około 16 dnia po owulacji endometrium nie ciężarnej macicy uwolni prostaglandynę F2alfa. PG F2alfa ma działanie luteolityczne, czyli rozpoczyna regresję ciałka żółtego.
Tabela 1. Hormony biorące udział w rozmnażaniu, miejsce ich powstania, główne funkcje i struktura chemiczna
Nazwa Miejsce powstania Główne funkcje Budowa chemiczna
Melatonina Szyszynka Wskaźnik długości dnia i nocy Indoloamina
GnRH Podwzgórze Stymuluje uwalnianie FSH i LH z przysadki Peptyd (10 aminokwasów)
FSH Przedni płat przysadki Samice:
Stymuluje wzrost i dojrzewanie pęcherzyka jajowego w jajnikach
Samce:
Stymuluje spermatogenezę Glikoproteina (>200 aminokwasów)
LH Przedni płat przysadki Samice:
Stymuluje dojrzewanie pęcherzyków jajnikowych, indukuje owulację, formowanie i utrzymywanie ciałka żółtego jajnika
Samce:
Stymuluje produkcję testosteronu Glikoproteina (>200 aminokwasów)
Estrogeny (17-beta-estradiol) Jajnik (komórki warstwy ziarnistej pęcherzyka jajnikowego) Indukuje zachowania rujowe. Stymuluje przedowulacyjny wylew GnRH Steroid
Inhibina Samice:
Jajnik (j.w.)
Samce:
Jądra (komórki Sertoliego) Hamuje uwalnianie FSH z przysadki (sprzężenie zwrotne) Peptyd
Progesteron Jajnik (ciałko żółte) Przygotowuje endometrium do zagnieżdżenia się zarodka. Podtrzymuje ciążę. Zmniejsza uwalnianie GnRH, co powstrzymuje nową owulację. Steroid
PGF2alfa Macica Regresja ciałka żółtego Kwas rozpuszczający tłuszcze
Luteolityczny mechanizm prostaglandyn nie jest całkowicie wyjaśniony, ale obejmuje on zmniejszenie ukrwienia ciałka żółtego przez obkurczenie naczyń oraz bezpośrednie działanie PG F2alfa na komórki lutealne. Oksytocyna produkowana w ciałku żółtym także odgrywa rolę w luteolizie. W rezultacie regresji ciałka żółtego obniża się stężenie progesteronu we krwi i ustępuje progesteronowy blok uwalniania GnRH z podwzgórza. Inicjuje to nową fazę pęcherzykową i ostateczne dojrzewanie pęcherzyka przedowulacyjnego. Okres dojrzewania pęcherzyka, rui i owulacji, charakteryzujący się produkcją estradiolu, jest nazywany fazą pęcherzykową cyklu. Faza dominacji progesteronu, od owulacji do luteolizy to faza lutealna.
Hormony wymienione w tym ustępie, które uczestniczą w rozrodzie, są przedstawione w tabeli 1. Wymienione są ich główne funkcje, pochodzenie i budowa chemiczna.
Należy zdawać sobie sprawę, że wymienione są tylko niektóre ze znanych działań hormonów, a nie wszystkie funkcje hormonów zostały poznane. Tabela uwzględnia jedynie znane funkcje endokrynne, ale większość hormonów ma także działanie parakrynowe, które jest słabo zbadane. Rozród u samic i u samców jest regulowany przez precyzyjne współgranie działań i odpowiedzi wywoływanych przez hormony. Chociaż w ciągu ostatniej dekady poczyniono znaczący postęp, całkowite zrozumienie tego niezwykle skomplikowanego mechanizmu jest nadal poza zasięgiem.
Regulacja rozrodu u samców
Zasady rozrodu u samców mają podobny wzór do występujących u samic. GnRH z podwzgórza stymuluje uwalnianie FSH i LH (u samców poprzednio zwane ICSH – interstitial cell stimulating hormone – hormon stymulujący komórki śródmiąższowe).
FSH działa na poziomie kanalików krętych jądra (komórki rozrodcze i komórki Sertoliego). Powoduje on stymulację spermatogenezy. Komórki Sertoliego produkują inhibinę, która wykazuje negatywne sprzężenie zwrotne na sekrecję FSH z podwzgórza.
LH stymuluje produkcję testosteronu przez komórki Leydiga. Testosteron wywiera wpływ na komórki Sertoliego i jest niezbędny dla udanej spermatogenezy. Ponadto, testosteron indukuje zmiany morfologiczne i typowo samcze zachowania. Testosteron wywiera ujemne sprzężenie zwrotne na uwalnianie LH poprzez zmniejszanie pulsacyjnego uwalniania GnRH z podwzgórza.
Podobny system kontroli rozrodu, ze stymulacją i sprzężeniami zwrotnymi, można spotkać u samic.
Sezonowość
W klimacie umiarkowanym wiele gatunków wykazuje sezonową aktywność płciową. Owce, kozy i konie są ich przykładem. Oznacza to, że okresy aktywności płciowej (okres oestrus) występują na przemian z okresami nieaktywności płciowej (okres anoestrus). Na przykład, u owiec aktywność płciowa rośnie przy skracaniu się długości dnia (gatunek dnia krótkiego), a u koni aktywność płciowa rośnie przy wydłużaniu się długości dnia (gatunek dnia długiego). Różnice momentu wystąpienia okresu płodnego u różnych gatunków wiążą się z różnicami w długości trwania ciąży. W klimacie umiarkowanym i zimnym zarówno konie jak i owce rodzą młode wiosną, w okresie obfitującym w pokarm, dając im najlepsze warunki do przetrwania.
Szyszynka jest głównym narządem odpowiedzialnym za regulację sezonowości. Poprzez oczy i zespół połączeń nerwowych długość dnia jest rejestrowana przez szyszynkę. Ilustruje to ryc. 5.
Szyszynka produkuje indoloaminy, z których najważniejszą jest melatonina. Melatonina jest produkowana i uwalniana w nocy (w ciemności). Kiedy zmniejsza się długość dnia wzrasta ekspozycja zwierzęcia na melatoninę. Wywiera to, w nie do końca poznany sposób, dodatni wpływ na sekrecje GnRH przez podwzgórze u gatunków krótkiego dnia, takich jak owca. U gatunków długiego dnia (konie) zwiększona ekspozycja na melatoninę w czasie długich nocy (krótkie dni) hamuje uwalnianie GnRH z podwzgórza. W ten sposób różnice w długości dnia są rejestrowane i przetwarzane w sygnały, które włączają bądź wyłączają aktywność płciową. Krowa straciła praktycznie swą sezonowość wskutek trwającego wieki procesu udomowiania
http://upload.miau.pl/20350.jpg
A teraz czekam na krytykę




{kind=link}